
Web alimentar marin - Marine food web

Comparativ cu mediile terestre, mediile marine au piramide de biomasă care sunt inversate la bază. În special, biomasa consumatorilor (copepode, krill, creveți, pești furajeri) este mai mare decât biomasa producătorilor primari . Acest lucru se întâmplă deoarece producătorii primari ai oceanului sunt mici fitoplancton care cresc și se reproduc rapid, astfel încât o masă mică poate avea o rată rapidă de producție primară . În schimb, mulți producători primari terestri semnificativi, cum ar fi pădurile mature , cresc și se reproduc lent, astfel încât este necesară o masă mult mai mare pentru a obține aceeași rată de producție primară.
Datorită acestei inversiuni, zooplanctonul este cel care formează majoritatea biomasei animale marine . Ca consumatori primari , zooplanctonul este legătura crucială dintre producătorii primari (în principal fitoplancton) și restul rețelei alimentare marine ( consumatori secundari ).
Dacă fitoplanctonul moare înainte de a fi mâncat, acesta coboară prin zona eufotică ca parte a zăpezii marine și se așează în adâncurile mării. În acest fel, fitoplanctonul sechestrează aproximativ 2 miliarde de tone de dioxid de carbon în ocean în fiecare an, determinând oceanul să devină o chiuvetă de dioxid de carbon care deține aproximativ 90% din tot carbonul sechestrat. Oceanul produce aproximativ jumătate din oxigenul din lume și stochează de 50 de ori mai mult dioxid de carbon decât atmosfera.
Un ecosistem nu poate fi înțeles fără cunoașterea modului în care rețeaua sa alimentară determină fluxul de materiale și energie. Fitoplanctonul produce autotrofic biomasă prin transformarea compușilor anorganici în compuși organici . În acest fel, fitoplanctonul funcționează ca baza rețelei alimentare marine prin susținerea tuturor celorlalte vieți din ocean. Al doilea proces central în rețeaua alimentară marină este bucla microbiană . Această buclă degradează bacteriile și arheele marine , remineralizează materia organică și anorganică și apoi reciclează produsele fie în rețeaua alimentară pelagică, fie prin depunerea lor ca sediment marin pe fundul mării.
Lanțuri alimentare și niveluri trofice
|
lanț alimentar marin (tipic) |
|---|
|
|
| ↓ |
| fitoplancton |
| ↓ |
| zooplancton erbivor |
| ↓ |
| zooplancton carnivor |
| ↓ |
|
|
| ↓ |
|
|
Pânzele alimentare sunt construite din lanțuri alimentare . Toate formele de viață din mare au potențialul de a deveni hrană pentru o altă formă de viață. În ocean, un lanț alimentar începe de obicei cu energia de la soare care alimentează fitoplanctonul și urmează un curs precum:
fitoplancton → zooplancton erbivor → zooplancton carnivor → alimentator de filtru → vertebrate de pradă
Fitoplanctonul nu are nevoie de alte organisme pentru hrană, deoarece au capacitatea de a-și produce propriile alimente direct din carbon anorganic, folosind lumina soarelui ca sursă de energie. Acest proces se numește fotosinteză și are ca rezultat ca fitoplanctonul să transforme carbonul natural în protoplasmă . Din acest motiv, se spune că fitoplanctonul este primul producător de la nivelul inferior sau al primului nivel al lanțului alimentar marin. Deoarece se află la primul nivel, se spune că au un nivel trofic de 1 (din greaca trophē care înseamnă mâncare). Fitoplanctonul este apoi consumat la următorul nivel trofic în lanțul trofic de animalele microscopice numite zooplancton.
Zooplanctonul constituie al doilea nivel trofic în lanțul trofic și include organisme microscopice unicelulare numite protozoare , precum și mici crustacee , cum ar fi copepodele și krilul , precum și larva peștilor, calmarilor, homarilor și crabilor. Organismele de la acest nivel pot fi considerate consumatori primari .
La rândul lor, zooplanktonul erbivor mai mic este consumat de zooplankterii carnivori mai mari, cum ar fi protozoarele și krilul mai mari de pradă , precum și de peștii furajeri , care sunt pești mici, de școală , care se hrănesc cu filtru . Aceasta reprezintă al treilea nivel trofic din lanțul alimentar.


Al patrulea nivel trofic este format din pești prădători , mamifere marine și păsări marine care consumă pești furajeri. Exemple sunt peștele-spadă , focile și sulurile .
Prădătorii de vârf, cum ar fi orcele , care pot consuma foci, și rechinii mako , care pot consuma pește-spadă, alcătuiesc un al cincilea nivel trofic. Balenele balene pot consuma zooplancton și krill direct, ducând la un lanț alimentar cu doar trei sau patru niveluri trofice.
În practică, nivelurile trofice nu sunt de obicei simple numere întregi, deoarece aceeași specie de consum se hrănește adesea în mai mult de un nivel trofic. De exemplu, o vertebrată marină mare poate mânca pești răpitori mai mici, dar poate mânca și hranitoare filtrante; de Stingray mănâncă crustaceele , dar hammerhead mananca atat crustacee si pisici de mare. Animalele se pot mânca și ele reciproc; de cod de cod mananca mai mici, precum și raci , și larve de cod Raci mânca. Obiceiurile de hrănire ale unui animal juvenil și, în consecință, nivelul său trofic, se pot schimba pe măsură ce crește.
Omul de știință în domeniul pescuitului, Daniel Pauly, stabilește valorile nivelurilor trofice la unul la producătorii primari și detritus , doi la erbivori și detritivori (consumatori primari), trei la consumatorii secundari, și așa mai departe. Definiția nivelului trofic, TL, pentru orice specie de consum este:

unde este nivelul fracțional trofic al prăzii j și reprezintă fracția de j din dieta i . În cazul ecosistemelor marine, nivelul trofic al majorității peștilor și al altor consumatori marini are valoare cuprinsă între 2,0 și 5,0. Valoarea superioară, 5.0, este neobișnuită, chiar și pentru peștii de talie mare, deși apare la prădătorii apex ai mamiferelor marine, cum ar fi urșii polari și balenele ucigașe. Ca punct de contrast, oamenii au un nivel trofic mediu de aproximativ 2,21, aproximativ la fel ca un porc sau o hamsie.


Prin taxon
Producători primari

La baza rețelei alimentare oceanice sunt alge unicelulare și alte organisme asemănătoare plantelor cunoscute sub numele de fitoplancton . Fitoplanctonul este un grup de autotrofe microscopice împărțite într-un ansamblu divers de grupuri taxonomice bazate pe morfologie , dimensiune și tip de pigment. Fitoplanctonul marin locuiește în mare parte în apele de suprafață iluminate de soare ca fotoautotrofe și necesită substanțe nutritive precum azotul și fosforul, precum și lumina soarelui pentru a fixa carbonul și a produce oxigen. Cu toate acestea, unii fitoplancton marin locuiesc în marea adâncă, adesea lângă gurile de adâncime , ca chimioautotrofe care utilizează surse de electroni anorganici, cum ar fi sulfura de hidrogen , fierul feros și amoniacul .
Fitoplanctonul marin constituie baza rețelei alimentare marine, reprezintă aproximativ jumătate din fixarea carbonului global și producția de oxigen prin fotosinteză și reprezintă o verigă cheie în ciclul global al carbonului. La fel ca plantele de pe uscat, fitoplanctonul folosește clorofila și alți pigmenți de recoltare a luminii pentru a efectua fotosinteza , absorbind dioxidul de carbon atmosferic pentru a produce zaharuri pentru combustibil. Clorofila din apă schimbă modul în care apa reflectă și absoarbe lumina soarelui, permițând oamenilor de știință să cartografieze cantitatea și locația fitoplanctonului. Aceste măsurători oferă oamenilor de știință informații valoroase asupra sănătății mediului oceanic și îi ajută pe oamenii de știință să studieze ciclul carbonului oceanic .

Printre fitoplancton se numără membrii dintr-un filum de bacterii numit cianobacterii . Cianobacteriile marine includ cele mai mici organisme fotosintetice cunoscute. Cel mai mic dintre toate, Prochlorococcus , are doar 0,5 până la 0,8 micrometri. În ceea ce privește numărul individual, Prochlorococcus este probabil cea mai abundentă specie de pe Pământ: un singur mililitru de apă de mare de suprafață poate conține 100.000 de celule sau mai mult. La nivel mondial, se estimează că există mai mulți octillioni (10 27 ) de indivizi. Proclorococul este omniprezent între 40 ° N și 40 ° S și domină în regiunile oligotrofe (nutrienți săraci) ale oceanelor. Bacteria reprezintă aproximativ 20% din oxigenul din atmosfera Pământului.
- Fitoplanctonul formează baza lanțului alimentar oceanic

Fitoplancton

Dinoflagelat

Diatomee



În oceane, cea mai mare producție primară este realizată de alge . Acesta este un contrast cu cel de pe uscat, unde cea mai mare producție primară este realizată de plante vasculare . Algele variază de la celule unice plutitoare la alge marine atașate , în timp ce plantele vasculare sunt reprezentate în ocean de grupuri precum ierburile marine și mangrovele . Producătorii mai mari, cum ar fi ierburile marine și algele marine , sunt în mare parte limitați la zona litorală și apele puțin adânci, unde se atașează la substratul subiacent și se află încă în zona fotică . Dar cea mai mare parte a producției primare de alge este realizată de fitoplancton.
Astfel, în mediile oceanice, primul nivel trofic de fund este ocupat în principal de fitoplancton , organisme microscopice în derivă, în mare parte alge unicelulare , care plutesc în mare. Majoritatea fitoplanctonului sunt prea mici pentru a fi văzute individual cu ochiul fără ajutor . Ele pot apărea ca o decolorare (adesea verde) a apei atunci când sunt prezente în număr suficient de mare. Deoarece își măresc biomasa în principal prin fotosinteză, ei trăiesc în stratul de suprafață luminat de soare ( zona eufotică ) a mării.
Cele mai importante grupuri de fitoplancton includ diatomeele și dinoflagelatele . Diatomeele sunt deosebit de importante în oceane, unde, potrivit unor estimări, contribuie până la 45% din producția primară totală a oceanului. Diatomeele sunt de obicei microscopice , deși unele specii pot ajunge până la 2 milimetri în lungime.
Consumatorii primari

Al doilea nivel trofic ( consumatorii primari ) este ocupat de zooplancton care se alimentează din fitoplancton . Împreună cu fitoplanctonul, formează baza piramidei alimentare care susține majoritatea marilor zone de pescuit ale lumii. Multe zooplancton sunt animale minuscule care se găsesc împreună cu fitoplanctonul în apele oceanice de suprafață și includ crustacee minuscule și larve și alevini de pește ( pești recent eclozați). Majoritatea zooplanctonului sunt filtratoare și folosesc anexe pentru a strecura fitoplanctonul în apă. Unele zooplancton mai mare se hrănesc și cu zooplancton mai mic. Unele zooplancton pot sări puțin pentru a evita prădătorii, dar nu pot înota cu adevărat. La fel ca fitoplanctonul, ele plutesc cu curenții, mareele și vânturile. Zooplanctoanele se pot reproduce rapid, populațiile lor pot crește până la treizeci la sută pe zi în condiții favorabile. Mulți duc o viață scurtă și productivă și ajung la maturitate rapid.
Cele oligotrichs sunt un grup de ciliate care au proeminent orale cilia aranjate ca un guler și rever. Acestea sunt foarte frecvente în comunitățile de plancton marin, de obicei găsite în concentrații de aproximativ unul pe mililitru. Sunt cei mai importanți erbivori din mare, prima verigă din lanțul trofic.
Alte grupuri deosebit de importante de zooplancton sunt copepodele și krilul . Copepodele sunt un grup de crustacei mici care se găsesc în habitatele din ocean și apă dulce . Sunt cea mai mare sursă de proteine din mare și sunt o pradă importantă pentru peștii furajeri. Krillul constituie următoarea sursă de proteine. Krilul este zooplancton de prădători deosebit de mare, care se hrănește cu zooplancton mai mic. Aceasta înseamnă că într-adevăr aparțin celui de-al treilea nivel trofic, consumatori secundari, împreună cu peștii furajeri.
- Zooplanctonul formează un al doilea nivel în lanțul alimentar oceanic

Vierme segmentat

Crustacee mici, asemănătoare creveților

Calmar planctonic juvenil




Împreună, fitoplanctonul și zooplanctonul alcătuiesc cea mai mare parte a planctonului din mare. Plancton este termenul aplicat oricăror organisme mici în derivă care plutesc în mare ( grecesc planktos = rătăcitor sau plutitor ). Prin definiție, organismele clasificate drept plancton nu pot înota împotriva curenților oceanici; nu pot rezista curentului ambiental și nu își pot controla poziția. În mediile oceanice, primele două niveluri trofice sunt ocupate în principal de plancton . Planctonul este împărțit în producători și consumatori. Producătorii sunt fitoplanctonul (grecul fiton = plantă), iar consumatorii, care mănâncă fitoplanctonul, sunt zooplanctonul (grecesc zoon = animal).
Meduzele sunt înotători încet și majoritatea speciilor fac parte din plancton. În mod tradițional meduze au fost văzute ca fundături trofice, jucătorii minore în trofice marine, organisme gelatinoasă cu un plan de organism în mare parte , pe baza de apa , care oferă o valoare puțin nutritivă sau de interes pentru alte organisme în afară de câteva prădători specializate , cum ar fi sunfish oceanului și broasca țestoasă Leatherback mare . Această viziune a fost recent contestată. Meduzele și, în general, zooplanctonul gelatinos, care includ săruri și ctenofori , sunt foarte diverse, fragile, fără părți dure, dificil de văzut și monitorizat, supuse unor fluctuații rapide ale populației și adesea trăiesc inconvenient departe de țărm sau adânc în ocean. Este dificil pentru oamenii de știință să detecteze și să analizeze meduzele în curajul prădătorilor, deoarece acestea se transformă în ciuperci atunci când sunt consumate și sunt digerate rapid. Dar meduzele înfloresc în număr mare și s-a arătat că formează componente majore în dietele de ton , suliță și pește-spadă , precum și diverse păsări și nevertebrate, cum ar fi caracatița , castraveții de mare , crabii și amfipodele . „În ciuda densității lor reduse de energie, contribuția meduzelor la bugetele energetice ale prădătorilor poate fi mult mai mare decât se presupune din cauza digestiei rapide, a costurilor reduse de captare, a disponibilității și a alimentării selective cu componentele mai bogate în energie. Hrănirea cu meduze poate face prădători marini susceptibili la ingestia de materiale plastice. "
Consumatori de ordin superior

- Nevertebrate marine
- Peşte
- Pești furajeri : peștii furajeri ocupă poziții centrale în rețelele alimentare oceanice . Organismele pe care le mănâncă sunt la un nivel trofic inferior , iar organismele care îl mănâncă se află la un nivel trofic superior. Peștii furajeri ocupă niveluri medii în rețeaua trofică, servind drept pradă dominantă pentru pești, păsări de mare și mamifere de nivel superior.
- Pești prădători
- Pește măcinat
- Alte vertebrate marine
În 2010, cercetătorii au descoperit că balenele transportă substanțe nutritive din adâncurile oceanului înapoi la suprafață, folosind un proces pe care l-au numit pompa pentru balene . Balenele se hrănesc la niveluri mai adânci în ocean, unde se găsește krilul , dar se întorc regulat la suprafață pentru a respira. Acolo balenele defecă un lichid bogat în azot și fier. În loc să se scufunde, lichidul rămâne la suprafața unde îl consumă fitoplanctonul . În Golful Maine, pompa pentru balene furnizează mai mult azot decât râurile.

Balenele cu cocoașă se aruncă de jos pentru a se hrăni cu pești furajeri

Sastele se scufundă de sus pentru a prinde pești furajeri

,_Belmont_-_geograph.org.uk_-_529175.jpg)
 Ciclul nutrienților pompei de balenă
Ciclul nutrienților pompei de balenă

Microorganisme

S-a recunoscut din ce în ce mai mult în ultimii ani că microorganismele marine joacă roluri mult mai mari în ecosistemele marine decât se credea anterior. Dezvoltările în metagenomică oferă cercetătorilor capacitatea de a dezvălui diversități ascunse anterior ale vieții microscopice, oferind un obiectiv puternic pentru vizualizarea lumii microbiene și potențialul de a revoluționa înțelegerea lumii vii. Tehnicile de analiză dietetică cu metabarcodare sunt utilizate pentru reconstituirea rețelelor alimentare la niveluri mai ridicate de rezoluție taxonomică și dezvăluie complexități mai profunde în rețeaua de interacțiuni.
Microorganismele joacă roluri cheie în rețelele alimentare marine. Calea de derivație virală este un mecanism care împiedică substanța organică particulă microbiană marină (POM) să migreze nivelurile trofice prin reciclarea acestora în materie organică dizolvată (DOM), care poate fi preluată cu ușurință de microorganisme. Manevrarea virală ajută la menținerea diversității în ecosistemul microbian prin împiedicarea unei singure specii de microbi marini să domine micro-mediul. DOM reciclat de calea de șunt viral este comparabil cu cantitatea generată de celelalte surse principale de DOM marin.
În general, carbonul organic dizolvat (DOC) este introdus în mediul oceanic din liza bacteriană, scurgerea sau exsudarea carbonului fix din fitoplancton (de exemplu, exopolimer mucilaginos din diatomee ), senescență bruscă a celulelor, hrănire neglijentă prin zooplancton, excreția deșeurilor produse de animale acvatice sau descompunerea sau dizolvarea particulelor organice din plante și soluri terestre. Bacteriile din bucla microbiană descompun acest detritus în particule pentru a utiliza această materie bogată în energie pentru creștere. Deoarece mai mult de 95% din materia organică din ecosistemele marine constă din compuși polimerici cu greutate moleculară mare (HMW) (de exemplu, proteine, polizaharide, lipide), doar o mică porțiune din materia organică dizolvată totală (DOM) este ușor utilizabilă pentru majoritatea marilor organisme la niveluri trofice superioare. Aceasta înseamnă că carbonul organic dizolvat nu este disponibil direct pentru majoritatea organismelor marine; bacteriile marine introduc acest carbon organic în rețeaua trofică, rezultând o energie suplimentară care devine disponibilă la niveluri trofice superioare.

așa cum a fost imaginat de un satelit în 2011



.jpg)


Un virus marin gigantic CroV infectează și provoacă moartea prin liza zooflagelatului marin Cafeteria roenbergensis . Acest lucru afectează ecologia de coastă, deoarece Cafeteria roenbergensis se hrănește cu bacterii găsite în apă. Atunci când există un număr redus de Cafeteria roenbergensis din cauza infecțiilor extinse cu CroV, populațiile bacteriene cresc exponențial. Impactul CroV asupra populațiilor naturale de C. roenbergensis rămâne necunoscut; cu toate acestea, sa constatat că virusul este foarte specific gazdei și nu infectează alte organisme strâns legate. Cafeteria roenbergensis este, de asemenea, infectată de un al doilea virus, virusul Mavirus , care este un virus satelit , ceea ce înseamnă că este capabil să se replice numai în prezența unui alt virus specific, în acest caz în prezența CroV. Acest virus interferează cu replicarea CroV, ceea ce duce la supraviețuirea celulelor C. roenbergensis . Mavirusul se poate integra în genomul celulelor C. roenbergensis și conferă astfel imunitate populației.
Ciuperci


După habitat
Pânze pelagice

Pentru ecosistemele pelagice, Legendre și Rassoulzadagan au propus în 1995 un continuum de căi trofice cu lanțul alimentar erbivor și bucla microbiană ca membri finali ai rețelei alimentare. Elementul liniar clasic al lanțului alimentar implică pășunatul de zooplancton pe fitoplancton mai mare și prădarea ulterioară pe zooplancton fie de zooplancton mai mare, fie de un alt prădător. Într-un astfel de lanț alimentar liniar, un prădător poate duce fie la biomasă de fitoplancton ridicată (într-un sistem cu fitoplancton, erbivor și prădător), fie la biomasă de fitoplancton redusă (într-un sistem cu patru niveluri). Modificările abundenței prădătorilor pot duce, astfel, la cascade trofice . Membrul final al buclei microbiene implică nu numai fitoplanctonul, ca resursă bazală, ci și carbon organic dizolvat . Carbonul organic dizolvat este utilizat de bacteriile heterotrofe pentru ca creșterea să fie predată de zooplanctonul mai mare. În consecință, carbonul organic dizolvat este transformat, printr-o buclă bacteriană-microzooplanctonică, în zooplancton. Aceste două căi de procesare a carbonului cu element final sunt conectate la mai multe niveluri. Fitoplanctonul mic poate fi consumat direct de microzooplancton.
Așa cum se ilustrează în diagrama din dreapta, carbonul organic dizolvat este produs în mai multe moduri și de diferite organisme, atât de producătorii primari, cât și de consumatorii de carbon organic. Eliberarea de DOC de către producătorii primari are loc pasiv prin scurgeri și activ în timpul creșterii dezechilibrate în timpul limitării nutrienților. O altă cale directă de la fitoplancton la bazinul organic dizolvat implică liza virală . Virușii marini sunt o cauză majoră a mortalității prin fitoplancton în ocean, în special în apele mai calde și cu latitudine mică. Hrănirea neglijentă de către erbivore și digestia incompletă a prăzii de către consumatori sunt alte surse de carbon organic dizolvat. Microbii heterotrofi folosesc enzime extracelulare pentru a solubiliza particulele de carbon organic și folosesc acesta și alte resurse de carbon organic dizolvat pentru creștere și întreținere. O parte din producția heterotrofă microbiană este utilizată de microzooplancton; o altă parte a comunității heterotrofe este supusă unei lize virale intense și acest lucru determină eliberarea din nou a carbonului organic dizolvat. Eficiența buclei microbiene depinde de mai mulți factori, dar în special de importanța relativă a prădării și a lizei virale pentru mortalitatea microbilor heterotrofi.
- Pânză alimentară pelagică

Rețeaua alimentară pelagică și pompa biologică. Legături între pompa biologică a oceanului și rețeaua trofică pelagică și capacitatea de a testa aceste componente de la distanță de pe nave, sateliți și vehicule autonome. Apele albastre deschis sunt zona eufotică , în timp ce apele albastre mai închise reprezintă zona crepusculului .

- Rețeaua alimentară mezopelagică

Impactul speciilor mezopelagice asupra bugetului global de carbon
DVM = migrație verticală diel NM = non-migrație


Gulile mezopelagice pot fi cele mai abundente vertebrate de pe planetă, deși se știe puțin despre ele.

Prădătorii gelatinoși ca acest nartedusan consumă cea mai mare diversitate de pradă mezopelagică



Oamenii de știință încep să exploreze mai detaliat zona crepusculară în mare parte necunoscută a mezopelagicului , cu o adâncime de 200 până la 1.000 de metri. Acest strat este responsabil pentru eliminarea a aproximativ 4 miliarde de tone de dioxid de carbon din atmosferă în fiecare an. Stratul mezopelagic este locuit de majoritatea biomasei peștelui marin .
Potrivit unui studiu din 2017, nartedusae consumă cea mai mare diversitate de pradă mezopelagică , urmată de sifonofori fizonecti , ctenofori și cefalopode . Importanța așa-numitei „pânze de jeleu” abia începe să fie înțeleasă, dar se pare că meduzele, ctenoforii și sifonoforii pot fi prădători cheie în rețelele alimentare pelagice profunde cu impact ecologic similar cu peștii de pradă și calamar. În mod tradițional, prădătorii gelatinoși erau considerați ineficienți ca furnizori de căi trofice marine, dar par să aibă roluri substanțiale și integrale în rețelele alimentare pelagice profunde. Migrația verticală Diel , un important mecanism activ de transport, permite mezozooplanctonului să sustragă dioxidul de carbon din atmosferă, precum și să furnizeze necesitățile de carbon pentru alte organisme mezopelagice.
Un studiu din 2020 a raportat că, până în 2050, încălzirea globală s-ar putea răspândi în oceanul adânc de șapte ori mai rapid decât este acum, chiar dacă emisiile de gaze cu efect de seră sunt reduse. Încălzirea în straturi mezopelagice și mai profunde ar putea avea consecințe majore pentru rețeaua hrănitoare oceanică profundă, deoarece speciile oceanice vor trebui să se deplaseze pentru a rămâne la temperaturi de supraviețuire.
- Peștii din amurg au aruncat o nouă lumină asupra ecosistemului oceanic Conversația , 10 februarie 2014.
- Un mister de ocean în trilioane The New York Times , 29 iunie 2015.
- Pești mesopelagici - expediție de circumnavigație Malaspina din 2010.

La suprafața oceanului

Habitatele de la suprafața oceanului stau la interfața dintre ocean și atmosferă. Biofilmelor-ca habitat la suprafața porturilor ocean microorganisme care trăiesc în suprafață, denumit în mod obișnuit ca neuston . Această vastă interfață aer-apă se află la intersecția proceselor majore de schimb aer-apă care acoperă mai mult de 70% din suprafața globală. Bacteriile din microstratul de suprafață al oceanului, numite bacterioneuston , sunt de interes datorită aplicațiilor practice precum schimbul de gaze cu efect de seră aer-mare, producerea de aerosoli marini activi din punct de vedere climatic și teledetecția oceanului. De un interes specific este producția și degradarea agenților tensioactivi (materiale active de suprafață) prin procese biochimice microbiene. Principalele surse de surfactanți din oceanul deschis includ fitoplanctonul, scurgerea terestră și depunerea din atmosferă.
Spre deosebire de înflorirea algelor colorate, bacteriile asociate agentului tensioactiv pot să nu fie vizibile în imagini color oceanice. Având capacitatea de a detecta aceste bacterii „invizibile” asociate agentului tensioactiv, folosind radar cu diafragmă sintetică, are beneficii imense în toate condițiile meteorologice, indiferent de nor, ceață sau lumina zilei. Acest lucru este deosebit de important în cazul vânturilor foarte puternice, deoarece acestea sunt condițiile în care au loc cele mai intense schimburi de gaze aer-mare și producția de aerosoli marini. Prin urmare, pe lângă imagini color prin satelit, imagini prin satelit SAR pot oferi informații suplimentare asupra unei imagini globale a proceselor biofizice la granița dintre ocean și atmosferă, schimburile de gaze cu efect de seră aer-mare și producția de aerosoli marini activi din punct de vedere climatic.
La fundul oceanului

Habitatele din fundul oceanului ( bentice ) stau la interfața dintre ocean și interiorul pământului.
- Seep și aerisiri

Pânze de coastă
Apele de coastă includ apele din estuare și peste platourile continentale . Acestea ocupă aproximativ 8% din suprafața oceanică totală și reprezintă aproximativ jumătate din întreaga productivitate a oceanului. Elementele nutritive cheie care determină eutrofizarea sunt azotul din apele de coastă și fosforul din lacuri. Ambele se găsesc în concentrații mari în guano (fecale de păsări marine), care acționează ca îngrășământ pentru oceanul înconjurător sau pentru un lac adiacent. Acidul uric este compusul dominant al azotului și în timpul mineralizării sale se produc diferite forme de azot.
Ecosistemele, chiar și cele cu frontiere aparent distincte, funcționează rareori independent de alte sisteme adiacente. Ecologiștii recunosc din ce în ce mai mult efectele importante pe care transportul de energie și substanțe nutritive între ecosisteme îl au asupra populațiilor și comunităților de plante și animale. Un exemplu bine cunoscut în acest sens este modul în care păsările marine concentrează substanțele nutritive derivate marine pe insulele de reproducere sub formă de fecale (guano) care conțin ~ 15-20% azot (N), precum și 10% fosfor. Acești nutrienți modifică dramatic funcționarea și dinamica ecosistemului terestru și pot sprijini creșterea productivității primare și secundare. Cu toate acestea, deși multe studii au demonstrat îmbogățirea cu azot a componentelor terestre datorită depunerii de guano în diferite grupuri taxonomice, doar câteva au studiat retroacțiunea sa asupra ecosistemelor marine și majoritatea acestor studii au fost limitate la regiuni temperate și ape cu nutrienți ridicați. La tropice, recifele de corali pot fi găsite adiacente insulelor cu populații mari de păsări marine care se reproduc și ar putea fi afectate potențial de îmbogățirea nutrienților locali datorită transportului de nutrienți derivați de păsările marine în apele înconjurătoare. Studiile asupra influenței guanoului asupra ecosistemelor marine tropicale sugerează că azotul din guano îmbogățește producătorii primari de apă de mare și recif.
Coralii de construcție de recif au nevoi esențiale de azot și, prosperând în apele tropicale sărace în nutrienți, unde azotul este un nutrient limitativ major pentru productivitatea primară, au dezvoltat adaptări specifice pentru conservarea acestui element. Stabilirea și întreținerea lor se datorează parțial simbiozei lor cu dinoflagelate unicelulare, Symbiodinium spp. (zooxanthellae), care poate prelua și reține azotul anorganic dizolvat (amoniu și azotat) din apele din jur. Aceste zooxantele pot recicla deșeurile animale și apoi le pot transfera înapoi la gazda coralului ca aminoacizi, amoniu sau uree. Coralii sunt, de asemenea, capabili să ingereze particule de sediment bogate în azot și plancton. Eutrofizarea litorală și aportul excesiv de nutrienți pot avea un impact puternic asupra coralilor, ducând la o scădere a creșterii scheletice,


În diagrama de mai sus din dreapta: (1) amonificarea produce NH3 și NH4 + și (2) nitrificarea produce NO3− prin oxidare NH4 +. (3) în condiții alcaline, tipice pentru fecalele păsărilor marine, NH3 se volatizează rapid și se transformă în NH4 +, (4) care este transportat în afara coloniei și prin depunere umedă exportat în ecosisteme îndepărtate, care sunt eutrofizate. Ciclul fosforului este mai simplu și are mobilitate redusă. Acest element se găsește într-o serie de forme chimice din materialul fecal al păsărilor marine, dar cel mai mobil și biodisponibil este ortofosfatul (5), care poate fi levigat de apele subterane sau superficiale.




pe un platou continental





Codarea în bare ADN poate fi utilizată pentru a construi structuri ale rețelei alimentare cu o rezoluție taxonomică mai bună la nodurile web. Aceasta oferă o identificare mai specifică a speciilor și o mai mare claritate cu privire exact la cine mănâncă pe cine. „Codurile de bare ADN și informațiile ADN pot permite noi abordări pentru construirea unor rețele mai mari de interacțiune și pot depăși unele obstacole în atingerea unei dimensiuni adecvate a eșantionului”.
O metodă recent aplicată pentru identificarea speciilor este metabarcodarea ADN-ului . Identificarea speciilor prin morfologie este relativ dificilă și necesită mult timp și expertiză. Metabarcodarea ADN-ului de secvențializare de mare viteză permite atribuirea taxonomică și, prin urmare, identificarea eșantionului complet cu privire la primerii specifici grupului aleși pentru amplificarea ADN anterioară .
- Codificare în bare ADN microbian
- Codarea în bare a ADN-ului algelor
- ADN-ul peștelui cod de bare
- Codificarea barei ADN în evaluarea dietei
- Păduri de alge
- Byrnes, JE, Reynolds, PL și Stachowicz, JJ (2007) „Invaziile și extincțiile remodelează rețelele alimentare costiere marine”. PLOS ONE , 2 (3): e295. doi : 10.1371 / journal.pone.0000295
Pânze polare

înconjurat de oceane

înconjurat de landmasses
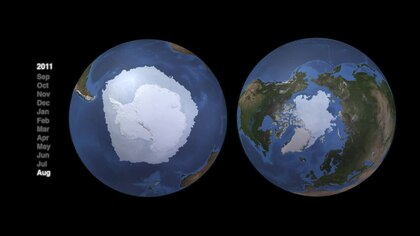
Sistemele marine arctice și antarctice au structuri topografice foarte diferite și, în consecință, au structuri ale rețelei alimentare foarte diferite. Atât rețelele alimentare pelagice arctice, cât și cele antarctice au fluxuri de energie caracteristice controlate în mare parte de câteva specii cheie. Dar nu există nici un singur web generic pentru nici unul. Căile alternative sunt importante pentru reziliență și menținerea fluxurilor de energie. Cu toate acestea, aceste alternative mai complicate asigură un flux de energie mai mic speciilor de nivel trofic superior. „Structura rețelei alimentare poate fi similară în diferite regiuni, dar speciile individuale care domină nivelurile trofice medii variază în funcție de regiunile polare”.



- Arctic


Rețeaua alimentară arctică este complexă. Pierderea gheții marine poate afecta în cele din urmă întreaga rețea trofică, de la alge și plancton la pești până la mamifere. Impactul schimbărilor climatice asupra unei anumite specii pot ondulație printr - o rețea de alimente și afectează o gamă largă de alte organisme ... Nu numai ca este declinul gheții marine afecta populațiile de urs polar , prin reducerea gradului de habitatului lor principal, este de asemenea afectându-le negativ prin intermediul efectelor rețelei alimentare. Scăderea duratei și întinderii gheții marine în Arctica duce la scăderea abundenței algelor de gheață, care prosperă în buzunare bogate în nutrienți în gheață. Aceste alge sunt consumate de zooplancton, care este la rândul său consumat de codul arctic, o sursă importantă de hrană pentru multe mamifere marine, inclusiv focile. Sigiliile sunt consumate de urșii polari. Prin urmare, scăderea algelor de gheață poate contribui la scăderea populației de urși polari.
În 2020 cercetătorii au raportat că măsurătorile din ultimele două decenii privind producția primară în Oceanul Arctic arată o creștere de aproape 60% din cauza concentrațiilor mai mari de fitoplancton . Ei fac ipoteza că noii nutrienți curg din alte oceane și sugerează că acest lucru înseamnă că oceanul arctic poate susține producția la nivel trofic mai ridicat și fixarea suplimentară a carbonului în viitor.




Săgeți gri: curgerea carbonului către heterotrofe
Săgeți verzi: căi majore ale fluxului de carbon către sau din mixotrofe
HCIL: ciliate heterotrofe ; MCIL: ciliate mixotrofe; HNF: nanoflagelate heterotrofe; DOC: carbon organic dizolvat; HDIN: dinoflagelați heterotrofi

- antarctic

Antarctica meduze Antarctica Diplulmaris sub gheata

Coloniile de alge Phaeocystis antarctica , un important fitoplankter al Mării Ross care domină începutul sezonului înflorește după ce gheața marină se retrage și exportă carbon semnificativ.

Pennate diatomee Fragilariopsis kerguelensis , găsit în întreaga Circumpolar curent Antarctica, este un factor cheie al globale pompei de silicat .

Un grup de balene ucigașe încearcă să disloceze un sigiliu de crabeater pe o floare de gheață
.jpg)




și mase de carbon (Gt C) în cutii întunecate



Fundație și specii cheie
.jpg)

.jpg)
Conceptul de specie de fundație a fost introdus în 1972 de Paul K. Dayton , care l-a aplicat anumitor membri ai comunităților de nevertebrate marine și alge . Din studiile efectuate în mai multe locații, a fost clar că există o mică mână de specii ale căror activități au avut un efect disproporționat asupra restului comunității marine și, prin urmare, acestea au fost cheia rezistenței comunității. Opinia lui Dayton era că concentrarea asupra speciilor de fundație ar permite o abordare simplificată pentru a înțelege mai rapid modul în care o comunitate în ansamblu ar reacționa la perturbări, cum ar fi poluarea, în loc să încerce sarcina extrem de dificilă de a urmări răspunsurile tuturor membrilor comunității simultan.
Speciile de fundație sunt specii care au un rol dominant în structurarea unei comunități ecologice , modelarea mediului său și definirea ecosistemului său. Astfel de ecosisteme sunt deseori numite după speciile de fundație, cum ar fi pajiști de iarbă de mare, straturi de stridii, recife de corali, păduri de alge și păduri de mangrove. De exemplu, mangrova roșie este o specie de bază comună în pădurile de mangrove. Rădăcina mangrovei oferă terenuri de pepiniere pentru peștii tineri, cum ar fi snapper . O specie de fundație poate ocupa orice nivel trofic într-o rețea trofică, dar tinde să fie un producător.


Conceptul speciei cheie a fost introdus în 1969 de zoologul Robert T. Paine . Paine a dezvoltat conceptul pentru a-și explica observațiile și experimentele privind relațiile dintre nevertebratele marine din zona intertidală (între liniile de mare și joasă), inclusiv stelele de mare și midiile . Unele stele de mare pradă pe arici de mare , midii și alte crustacee , care nu au alte prădătorilor naturali. Dacă steaua de mare este scoasă din ecosistem, populația de midii explodează necontrolat, alungând majoritatea celorlalte specii.
Speciile Keystone sunt specii care au efecte mari, disproporționate față de numărul lor, în rețelele alimentare ecosistemice. Un ecosistem poate experimenta o schimbare dramatică dacă o specie cheie este eliminată, chiar dacă specia respectivă era o mică parte a ecosistemului prin măsuri de biomasă sau productivitate . Vidrele de mare limita daunele aricii de mare aplicate padurilor de kelp . Când vidrele de mare de pe coasta de vest a Americii de Nord au fost vânate comercial pentru blana lor, numărul lor a scăzut la niveluri atât de scăzute încât nu au putut controla populația de arici de mare. La rândul lor, puii pășeau atât de puternic în țărmurile de alge, încât pădurile de alge au dispărut în mare măsură, împreună cu toate speciile care depindeau de ele. Reintroducerea vidrelor de mare a permis restaurarea ecosistemului de vară.
Poziția topologică
Rețelele de interacțiuni trofice pot furniza o mulțime de informații despre funcționarea ecosistemelor marine. Dincolo de obiceiurile de hrănire, trei trăsături suplimentare (mobilitate, dimensiune și habitat) ale diferitelor organisme pot completa această viziune trofică.

Pentru a susține buna funcționare a ecosistemelor, este nevoie să înțelegem mai bine întrebarea simplă adresată de Lawton în 1994: Ce fac speciile în ecosisteme? Întrucât rolurile ecologice și pozițiile rețelei alimentare nu sunt independente, trebuie pusă întrebarea despre ce tip de specii ocupă diferite poziții din rețea. Încă de la primele încercări de identificare a speciilor cheie, a existat un interes pentru locul lor în rețelele alimentare. Mai întâi li s-a sugerat că ar fi fost prădători de top, apoi și plante, erbivore și paraziți. Atât pentru ecologia comunității, cât și pentru biologia conservării, ar fi util să știm unde se află în rețele trofice complexe.
Un exemplu al acestui tip de analiză a rețelei este prezentat în diagramă, pe baza datelor dintr-o rețea alimentară marină. Arată relațiile dintre pozițiile topologice ale nodurilor web și valorile de mobilitate ale organismului implicat. Nodurile web sunt codificate în formă în funcție de mobilitate și codificate în culori folosind indici care subliniază (A) grupurile de jos în sus (sesile și derivați) și grupurile (B) din partea de sus a rețelei alimentare.
Importanța relativă a organismelor variază în funcție de timp și spațiu, iar examinarea bazelor de date mari poate oferi informații generale asupra problemei. Dacă diferite tipuri de organisme ocupă diferite tipuri de poziții în rețea, atunci ajustarea pentru acest lucru în modelarea rețelei alimentare va avea ca rezultat predicții mai fiabile. Comparațiile indicilor de centralitate între ei (asemănarea gradului de centralitate și centralitate a apropierii , indicii cheie și keystoneness și indicii de centralitate față de nivelul trofic (majoritatea speciilor cu centralitate ridicată la niveluri trofice medii) au fost făcute pentru a înțelege mai bine pozițiile importante ale organismelor în rețelele alimentare. Extinderea acestui interes prin adăugarea de date despre trăsături la grupurile trofice ajută la interpretarea biologică a rezultatelor. Relațiile dintre indicii de centralitate au fost studiate și pentru alte tipuri de rețele, inclusiv rețelele de habitate. Cu baze de date mari și noi analize statistice, întrebări ca acestea pot fi reinvestigate și cunoștințele pot fi actualizate.
Interacțiuni criptice

Interacțiunile criptice, interacțiuni care sunt „ascunse la vedere”, apar în întreaga rețea planctonică marină, dar în prezent sunt în mare măsură trecute cu vederea prin metode stabilite, ceea ce înseamnă că colectarea datelor pe scară largă pentru aceste interacțiuni este limitată. În ciuda acestui fapt, dovezile actuale sugerează că unele dintre aceste interacțiuni pot avea impacturi perceptibile asupra dinamicii web și a rezultatelor modelului. Incorporarea interacțiunilor criptice în modele este deosebit de importantă pentru acele interacțiuni care implică transportul de nutrienți sau energie.
Diagrama ilustrează fluxurile de materiale, populațiile și bazinele moleculare care sunt afectate de cinci interacțiuni criptice: mixotrofie , diferențe ontogenetice și specii, hrănire microbiană, auxotrofie și partiționare celulară a carbonului. Aceste interacțiuni pot avea efecte sinergice pe măsură ce regiunile rețelei alimentare pe care le afectează se suprapun. De exemplu, partiția carbonului celular din fitoplancton poate afecta atât bazinele de materie organică din aval utilizate în alimentarea încrucișată microbiană și schimbate în caz de auxotrofie, cât și selecția prăzilor bazate pe diferențe ontogenetice și specii.
Simplificări precum „zooplanctonul consumă fitoplancton”, „fitoplanctonul preia nutrienți anorganici”, „producția primară brută determină cantitatea de carbon disponibilă pentru rețeaua trofică” etc., i-au ajutat pe oamenii de știință să explice și să modeleze interacțiunile generale din mediul acvatic. Metodele tradiționale s-au concentrat pe cuantificarea și calificarea acestor generalizări, dar progresele rapide în genomică, limitele de detectare a senzorilor, metodele experimentale și alte tehnologii din ultimii ani au arătat că generalizarea interacțiunilor în cadrul comunității planctonice poate fi prea simplă. Aceste îmbunătățiri ale tehnologiei au expus o serie de interacțiuni care par a fi criptice, deoarece eforturile de eșantionare în bloc și metodele experimentale sunt părtinitoare împotriva lor.
Complexitate și stabilitate

Pânzele alimentare oferă un cadru în cadrul căruia poate fi organizată o rețea complexă de interacțiuni prădător-pradă. Un model de rețea alimentară este o rețea de lanțuri alimentare . Fiecare lanț alimentar începe cu un producător primar sau autotrof , un organism, cum ar fi o algă sau o plantă, care este capabilă să-și fabrice propria hrană. Următorul lanț este un organism care se hrănește cu producătorul primar, iar lanțul continuă în acest fel ca un șir de prădători succesivi. Organismele din fiecare lanț sunt grupate în niveluri trofice , în funcție de câte verigi sunt eliminate de la producătorii primari. Lungimea lanțului sau nivelul trofic este o măsură a numărului de specii întâlnite pe măsură ce energia sau nutrienții se deplasează de la plante la prădători de vârf. Energia alimentară curge de la un organism la altul și la următorul și așa mai departe, cu o anumită energie pierdută la fiecare nivel. La un nivel trofic dat poate exista o specie sau un grup de specii cu aceiași prădători și pradă.
În 1927, Charles Elton a publicat o sinteză influentă asupra utilizării rețelelor alimentare, care a dus la transformarea acestora într-un concept central în ecologie. În 1966, interesul pentru rețelele alimentare a crescut după studiul experimental și descriptiv realizat de Robert Paine asupra țărmurilor intertidale, sugerând că complexitatea rețelei alimentare a fost esențială pentru menținerea diversității speciilor și a stabilității ecologice. Mulți ecologiști teoretici, inclusiv Robert May și Stuart Pimm , au fost determinați de această descoperire și de alții să examineze proprietățile matematice ale rețelelor alimentare. Conform analizelor lor, rețelele alimentare complexe ar trebui să fie mai puțin stabile decât rețelele alimentare simple. Aparentul paradox dintre complexitatea rețelelor alimentare observate în natură și fragilitatea matematică a modelelor rețelei alimentare este în prezent o zonă de studiu intens și dezbatere. Paradoxul se poate datora parțial diferențelor conceptuale dintre persistența unei rețele alimentare și stabilitatea echilibrată a unei rețele alimentare.
O cascadă trofică poate apărea într-o rețea alimentară dacă este suprimat un nivel trofic în rețea.
De exemplu, o cascadă de sus în jos poate apărea dacă prădătorii sunt suficient de eficienți în prădare pentru a reduce abundența sau pentru a modifica comportamentul prăzii lor , eliberând astfel următorul nivel trofic inferior din prădare. O cascadă de sus în jos este o cascadă trofică în care consumatorul / prădătorul de top controlează populația de consumatori primari . La rândul său, populația producătoare primară prosperă. Eliminarea prădătorului superior poate modifica dinamica rețelei alimentare. În acest caz, consumatorii primari ar suprapopula și exploata producătorii primari. În cele din urmă, nu ar exista suficienți producători primari pentru a susține populația consumatoare. Stabilitatea top-down a pânzei alimentare depinde de concurență și prădare la nivelurile trofice superioare. De asemenea, speciile invazive pot modifica această cascadă prin îndepărtarea sau devenirea unui prădător de vârf. Este posibil ca această interacțiune să nu fie întotdeauna negativă. Studiile au arătat că anumite specii invazive au început să schimbe cascadele; și, în consecință, degradarea ecosistemului a fost reparată. Un exemplu de cascadă într-un ecosistem complex, deschis, a avut loc în nord-vestul Atlanticului în anii 1980 și 1990. Îndepărtarea codului Atlantic ( Gadus morhua ) și a altor pești de sol prin pescuitul excesiv susținut a dus la creșterea abundenței speciilor de pradă pentru acești pești de sol, în special pești furajeri mai mici și nevertebrate, cum ar fi crabul de zăpadă nordic ( Chionoecetes opilio ) și creveții nordici ( Pandalus borealis ). Abundența crescută a acestor specii de pradă a modificat comunitatea de zooplancton care servește ca hrană pentru peștii mai mici și nevertebrate ca efect indirect. Cascadele de sus în jos pot fi importante pentru a înțelege efectele lovite ale eliminării prădătorilor de top din rețelele alimentare, așa cum au făcut oamenii în multe locuri prin vânătoare și pescuit .
Într-o cascadă de jos în sus, populația producătorilor primari va controla întotdeauna creșterea / scăderea energiei la nivelurile trofice superioare. Producătorii primari sunt plantele, fitoplanctonul și zooplanctonul care necesită fotosinteză. Deși lumina este importantă, populațiile primare producătoare sunt modificate de cantitatea de nutrienți din sistem. Această rețea alimentară se bazează pe disponibilitatea și limitarea resurselor. Toate populațiile vor avea creștere dacă există inițial o cantitate mare de nutrienți.
Comparații terestre

Comparativ cu piramidele de biomasă terestră, piramidele acvatice sunt în general inversate la bază
.jpg)
.jpg)
Mediile marine pot avea inversiuni în piramidele lor de biomasă. În special, biomasa consumatorilor (copepode, krill, creveți, pești furajeri) este, în general, mai mare decât biomasa producătorilor primari. Acest lucru se întâmplă deoarece producătorii primari ai oceanului sunt în mare parte fitoplancton mic, care au trăsături de strateg r de creștere și reproducere rapidă, astfel încât o masă mică poate avea o rată rapidă de producție primară. În schimb, mulți producători primari terestri, cum ar fi pădurile mature, au trăsături de strateg K de creștere și reproducere lentă, astfel încât este necesară o masă mult mai mare pentru a obține aceeași rată de producție primară. Rata de producție împărțită la cantitatea medie de biomasă care o atinge este cunoscută ca raportul Producție / Biomasă (P / B) al unui organism. Producția este măsurată în funcție de cantitatea de mișcare a masei sau a energiei pe zonă pe unitate de timp. În schimb, măsurarea biomasei se face în unități de masă pe unitate de suprafață sau volum. Raportul P / B utilizează unități de timp invers (exemplu: 1 / lună). Acest raport permite o estimare a cantității de flux de energie comparativ cu cantitatea de biomasă la un nivel trofic dat, permițând efectuarea de delimitări între nivelurile trofice. Raportul P / B scade cel mai frecvent pe măsură ce crește nivelul trofic și mărimea organismului, cu organisme mici, efemere, care conțin un raport P / B mai mare decât cele mari, de lungă durată.
Exemple: pinul bristlecone poate trăi mii de ani și are un raport foarte mic de producție / biomasă. Cianobacteria Prochlorococcus trăiește aproximativ 24 de ore și are un raport foarte mare de producție / biomasă.
În oceane, cea mai mare producție primară este realizată de alge . Acesta este un contrast cu cel de pe uscat, unde cea mai mare producție primară este realizată de plante vasculare .
|
Comparația productivității în ecosistemele marine și terestre
|
|||
|---|---|---|---|
| Ecosistem | Productivitate primară netă miliarde de tone pe an |
Total biomasă vegetală miliarde de tone |
Cifra de afaceri de ani |
|
Marin
|
45–55
|
1-2
|
0,02-0,06
|
|
Terestru
|
55-70
|
600–1000
|
9–20
|

Producătorilor acvatici, cum ar fi algele planctonice sau plantele acvatice, le lipsește acumularea mare de creștere secundară care există în copacii lemnoși ai ecosistemelor terestre. Cu toate acestea, sunt capabili să se reproducă suficient de repede pentru a susține o biomasă mai mare de pășunători. Aceasta inversează piramida. Consumatorii primari au o durată de viață mai lungă și rate de creștere mai lente, care acumulează mai multă biomasă decât producătorii pe care îi consumă. Fitoplanctonul trăiește doar câteva zile, în timp ce zooplanctonul care mănâncă fitoplanctonul trăiește câteva săptămâni, iar peștii care mănâncă zooplanctonul trăiesc câțiva ani consecutivi. De asemenea, prădătorii acvatici tind să aibă o rată a mortalității mai mică decât consumatorii mai mici, ceea ce contribuie la modelul piramidal inversat. Structura populației, ratele de migrație și refugiul de mediu pentru pradă sunt alte cauze posibile pentru piramidele cu biomasă inversată. Cu toate acestea, piramidele energetice vor avea întotdeauna o formă de piramidă verticală dacă sunt incluse toate sursele de energie alimentară, deoarece aceasta este dictată de a doua lege a termodinamicii . "
Cea mai mare parte a materiei organice produse este consumată în cele din urmă și respirată de carbon anorganic . Rata la care materia organică este păstrată prin înmormântare prin acumularea de sedimente este de numai 0,2 și 0,4 miliarde de tone pe an, reprezentând o fracțiune foarte mică din producția totală. Producția globală de fitoplancton este de aproximativ 50 de miliarde de tone pe an, iar biomasa de fitoplancton este de aproximativ un miliard de tone, ceea ce implică un timp de rulare de o săptămână. Macrofitele marine au o biomasă globală similară, dar o producție de doar un miliard de tone pe an, ceea ce implică o perioadă de rulare de un an. Aceste rate mari de rotație (comparativ cu rotația globală a vegetației terestre de una până la două decenii) implică nu numai producția constantă, ci și consumul eficient de materie organică. Există mai multe căi de pierdere a materiei organice (respirația prin autotrofi și heterotrofi, pășunat, liză virală, cale detritică), dar toate au ca rezultat respirația și eliberarea carbonului anorganic.
.jpeg)
Efecte antropice

- Pescuitul excesiv
- Acidificare
Pteropodele și stelele fragile formează împreună baza rețelelor alimentare arctice și ambele sunt grav deteriorate de acidificare. Cojile pteropode se dizolvă odată cu acidificarea crescândă și stelele fragile pierd masa musculară atunci când re-cresc apendicele. În plus, ouăle stelei fragile mor în câteva zile când sunt expuse condițiilor așteptate rezultate din acidificarea arctică. Acidificarea amenință să distrugă rețelele alimentare arctice de la bază în sus. Apele arctice se schimbă rapid și sunt avansate în procesul de a fi insaturate cu aragonit. Pânzele alimentare arctice sunt considerate simple, ceea ce înseamnă că există câțiva pași în lanțul alimentar de la organisme mici la prădători mai mari. De exemplu, pteropodele sunt „un element de pradă cheie al unui număr de prădători superiori - plancton mai mare, pești, păsări marine, balene”.
.jpg)

- Schimbarea climei
„Rezultatele noastre arată cum schimbările climatice viitoare pot slăbi rețelele alimentare marine prin flux redus de energie la niveluri trofice mai ridicate și o trecere la un sistem mai bazat pe detritus, ducând la simplificarea rețelei alimentare și la modificarea dinamicii producător-consumator, ambele având o importanță importantă implicații pentru structurarea comunităților bentice. "
„... temperaturile crescute reduc fluxul vital de energie de la producătorii de alimente primare din partea de jos (de exemplu, alge), către consumatorii intermediari (erbivori), până la prădătorii din partea superioară a rețelelor alimentare marine. Astfel de perturbări în transferul de energie pot duce potențial la o scădere a disponibilității hranei pentru prădătorii de top, care, la rândul său, poate duce la impacturi negative pentru multe specii marine din aceste rețele alimentare ... "În timp ce schimbările climatice au crescut productivitatea plantelor, acest lucru s-a datorat în principal unei expansiuni a cianobacteriilor ( mici alge-albastre-verzi) ", a spus domnul Ullah." Această productivitate primară crescută nu susține rețelele alimentare, totuși, deoarece aceste cianobacterii sunt în mare parte neplăcute și nu sunt consumate de erbivore. Înțelegerea modului în care funcționează ecosistemele sub efectele încălzirii globale este o provocare în cercetarea ecologică. Majoritatea cercetărilor privind încălzirea oceanelor implică experimente simplificate, pe termen scurt, bazate doar pe una sau câteva specii. "

