
Producție primară marină - Marine primary production

Producția primară marină este sinteza chimică în ocean a compușilor organici din dioxid de carbon atmosferic sau dizolvat . Apare în principal prin procesul de fotosinteză , care folosește lumina ca sursă de energie, dar apare și prin chemosinteză , care folosește oxidarea sau reducerea compușilor chimici anorganici ca sursă de energie. Aproape toată viața de pe Pământ se bazează direct sau indirect pe producția primară . Organismele responsabile de producția primară sunt numite producători primari sau autotrofe .
Majoritatea producției primare marine este generată de o colecție diversă de microorganisme marine numite alge și cianobacterii . Împreună, aceștia formează principalii producători primari de la baza lanțului alimentar oceanic și produc jumătate din oxigenul lumii. Producătorii primari marini susțin aproape toată viața animalelor marine, generând aproape tot oxigenul și hrana pentru care animalele marine trebuie să existe. Unii producători primari marini sunt, de asemenea , ingineri ecosistemici care schimbă mediul și oferă habitate pentru alte vieți marine.
Producția primară în ocean poate fi contrastată cu producția primară pe uscat. La nivel global oceanul și terenul produc fiecare aproximativ aceeași cantitate de producție primară, dar în ocean producția primară provine în principal din cianobacterii și alge, în timp ce pe uscat provine în principal din plante vasculare .
Algele marine include în mare parte invizibilă și de multe ori unicelular microalge , care , împreună cu cianobacterii formează oceanul fitoplanctonului , precum și mai mare, mai vizibile si complexe multicelular macroalgele frecvent numit alge marine . Algele marine se găsesc de-a lungul zonelor de coastă, trăind pe podeaua rafturilor continentale și spălate în zonele intertidale . Unele alge marine plutesc cu plancton în apele de suprafață luminate de soare ( zona epipelagică ) ale oceanului deschis.
Înapoi în Silurian , unele fitoplancton au evoluat în alge roșii , maronii și verzi . Aceste alge au invadat apoi pământul și au început să evolueze în plantele terestre pe care le cunoaștem astăzi. Mai târziu, în Cretacic, unele dintre aceste plante terestre s-au întors la mare ca mangrove și ierburi marine . Acestea se găsesc de-a lungul coastelor în regiunile intertidale și în apa sălbatică a estuarelor . În plus, unele ierburi marine, cum ar fi algele marine, pot fi găsite la adâncimi de până la 50 de metri, atât pe fundul moale, cât și pe cel dur al platoului continental.
Producători primari marini

• Roșu = diatomee (fitoplancton mare, care necesită silice)
• Galben = flagelați (alt fitoplancton mare)
• Verde = proclorococ (fitoplancton mic care nu poate folosi azotat)
• Cian = sinechococ (alt fitoplancton mic)
Opacitatea indică concentrația biomasei de carbon. În special, rolul vârtejurilor și filamentelor ( caracteristici mezoscale ) par important în menținerea biodiversității ridicate în ocean.
| Parte a unei serii pe |
| Ciclul carbonului |
|---|
 |
Producătorii primari sunt organismele autotrofe care își fac propria hrană în loc să mănânce alte organisme. Aceasta înseamnă că producătorii primari devin punctul de plecare în lanțul alimentar pentru organismele heterotrofe care mănâncă alte organisme. Unii producători primari marini sunt bacterii și arhee specializate, care sunt chemotrofe , producându-și propria hrană prin adunarea în jurul orificiilor hidrotermale și a scurgerilor reci și folosind chemosinteza . Cu toate acestea, cea mai mare parte a producției primare marine provine din organisme care utilizează fotosinteza dioxidului de carbon dizolvat în apă. Acest proces folosește energia din lumina soarelui pentru a transforma apa și dioxidul de carbon în zaharuri care pot fi utilizate atât ca sursă de energie chimică, cât și a moleculelor organice care sunt utilizate în componentele structurale ale celulelor. Producătorii primari marini sunt importanți, deoarece susțin aproape toată viața animalelor marine, generând cea mai mare parte a oxigenului și a alimentelor care furnizează altor organisme energia chimică de care au nevoie pentru a exista.
Principalii producători marini primari sunt cianobacteriile , algele și plantele marine. Oxigenul eliberat ca un produs secundar al fotosintezei este nevoie de aproape toate lucrurile pentru a efectua trai respirația celulară . În plus, producătorii primari sunt influenți în ciclurile globale de carbon și apă . Stabilizează zonele de coastă și pot oferi habitate animalelor marine. Termenul diviziune a fost folosit în mod tradițional în loc de filum atunci când se discută despre producătorii primari, deși Codul internațional de nomenclatură pentru alge, ciuperci și plante acceptă acum termenii ca echivalenți.
Într-o inversare a tiparului pe uscat, în oceane, aproape toată fotosinteza este realizată de alge și cianobacterii, cu o mică parte contribuită de plantele vasculare și alte grupuri. Algele cuprind o gamă variată de organisme, variind de la celule plutitoare simple până la alge marine atașate . Acestea includ fotoautotrofe dintr-o varietate de grupuri. Eubacteriile sunt fotosintetizatoare importante atât în ecosistemele oceanice, cât și pe cele terestre și, în timp ce unele arhee sunt fototrofe , nu se știe că utilizează fotosinteza care evoluează cu oxigen. Un număr de eucariote contribuie semnificativ la producția primară în ocean, inclusiv alge verzi , alge brune și alge roșii și un grup divers de grupuri unicelulare. Plantele vasculare sunt, de asemenea, reprezentate în ocean de grupuri precum ierburile marine .
Spre deosebire de ecosistemele terestre, majoritatea producției primare în ocean este realizată de organisme microscopice cu viață liberă numite fitoplancton . S-a estimat că jumătate din oxigenul din lume este produs de fitoplancton. Autotrofe mai mari, cum ar fi iarbele marine și macroalge ( alge marine ) sunt, în general, limitate la zona litorală și apele adânci adânci, unde se pot atașa la substratul subiacent, dar se află încă în zona fotică . Există excepții, cum ar fi Sargassum , dar marea majoritate a producției cu plutire liberă are loc în cadrul organismelor microscopice.
Factorii care limitează producția primară în ocean sunt, de asemenea, foarte diferiți de cei de pe uscat. Disponibilitatea apei, evident, nu este o problemă (deși salinitatea ei poate fi). În mod similar, temperatura, deși afectează ratele metabolice (vezi Q 10 ), variază mai puțin în ocean decât pe uscat, deoarece capacitatea de căldură a apei de mare tamponează temperatura, iar formarea gheții marine o izolează la temperaturi mai scăzute. Cu toate acestea, disponibilitatea luminii, sursa de energie pentru fotosinteză și a nutrienților minerali , elemente de bază pentru o nouă creștere, joacă un rol crucial în reglarea producției primare în ocean. Modelele disponibile ale Sistemului Pământ sugerează că modificările bio-geochimice oceanice în curs ar putea declanșa reduceri ale centralei nucleare oceanice între 3% și 10% din valorile actuale, în funcție de scenariul de emisii.
În 2020 cercetătorii au raportat că măsurătorile din ultimele două decenii ale producției primare în Oceanul Arctic arată o creștere de aproape 60% din cauza concentrațiilor mai mari de fitoplancton . Ei presupun că noii nutrienți curg din alte oceane și sugerează că acest lucru înseamnă că oceanul arctic ar putea să susțină producția la nivel trofic mai ridicat și fixarea suplimentară a carbonului în viitor.
Cianobacterii

Cianobacteriile sunt un filum (diviziune) de bacterii, variind de la unicelulare la filamentoase și incluzând specii coloniale , care fixează carbonul anorganic în compuși organici de carbon. Se găsesc aproape peste tot pe pământ: în solul umed, atât în mediul de apă dulce, cât și în cel marin, și chiar pe rocile din Antarctica. În special, unele specii apar ca celule în derivă care plutesc în ocean și, ca atare, au fost printre primele fitoplancton . Aceste bacterii funcționează ca algele prin faptul că pot procesa azotul din atmosferă atunci când niciuna nu se află în ocean.

Primii producători primari care au folosit fotosinteza au fost cianobacteriile oceanice în urmă cu aproximativ 2,3 miliarde de ani. Eliberarea de oxigen molecular de către cianobacterii ca produs secundar al fotosintezei a indus schimbări globale în mediul Pământului. Deoarece oxigenul era toxic pentru majoritatea vieților de pe Pământ la acea vreme, acest lucru a dus la aproape dispariția organismelor intolerante la oxigen , o schimbare dramatică care a redirecționat evoluția principalelor specii de animale și plante.
.jpg)
Micul cianobacteriu marin Prochlorococcus , descoperit în 1986, face astăzi parte din baza lanțului alimentar oceanic și reprezintă mai mult de jumătate din fotosinteza oceanului deschis și aproximativ 20% din oxigenul din atmosfera Pământului. Este probabil cel mai abundent gen de pe Pământ: un singur mililitru de apă de mare de suprafață poate conține 100.000 de celule sau mai mult.
Inițial, biologii au crezut că cianobacteriile sunt alge și au denumit-o „alge albastru-verzi”. Opinia mai recentă este că cianobacteriile sunt bacterii și, prin urmare, nici măcar nu se află în același Regat ca algele. Majoritatea autorităților exclud toate procariotele și, prin urmare, cianobacteriile din definiția algelor.

Pigmenți biologici
Pigmenții biologici sunt orice material colorat din celulele vegetale sau animale. Toți pigmenții biologici absorb în mod selectiv anumite lungimi de undă ale luminii în timp ce reflectă altele. Funcția principală a pigmenților din plante este fotosinteza , care folosește pigmentul verde clorofilă și mai mulți pigmenți coloranți care absorb cât mai multă energie luminoasă posibil. Clorofila este pigmentul primar la plante; este un clor care absoarbe lungimile de undă galbene și albastre ale luminii în timp ce reflectă verde . Prezența și abundența relativă a clorofilei conferă plantelor culoarea lor verde. Algele verzi și plantele posedă două forme ale acestui pigment: clorofila a și clorofila b . Kelps , diatomee și alte heterokonturi fotosintetice conțin clorofilă c în loc de b , în timp ce algele roșii posedă doar clorofilă a . Toate clorofilele servesc ca mijloace primare folosite de plante pentru a intercepta lumina pentru a alimenta fotosinteza.
Cloroplastele
.jpg)

Cloroplastele (din grecescul chloros pentru verde și plasturile pentru „cel care se formează”) sunt organite care conduc fotosinteza , unde pigmentul fotosintetic clorofilă captează energia din lumina soarelui , o convertește și o stochează în moleculele de stocare a energiei în timp ce eliberează oxigen din apa din celulele vegetale si algale . Apoi folosesc energia stocată pentru a produce molecule organice din dioxid de carbon într-un proces cunoscut sub numele de ciclul Calvin .
Un cloroplast este un tip de organet cunoscut sub numele de plastid , caracterizat prin cele două membrane și o concentrație ridicată de clorofilă . Sunt foarte dinamici - circulă și se deplasează în interiorul celulelor vegetale și, uneori, se ciupesc în două pentru a se reproduce. Comportamentul lor este puternic influențat de factori de mediu precum culoarea și intensitatea luminii. Cloroplastele, la fel ca mitocondriile , conțin propriul ADN , despre care se crede că este moștenit de la strămoșul lor - o cianobacterie fotosintetică care a fost înghițită de o celulă eucariotă timpurie . Cloroplastele nu pot fi produse de celula vegetală și trebuie moștenite de fiecare celulă fiică în timpul diviziunii celulare.
Majoritatea cloroplastelor pot fi urmărite probabil într-un singur eveniment endosimbiotic , când o cianobacterie a fost înghițită de eucariot. În ciuda acestui fapt, cloroplastele pot fi găsite într-un set extrem de larg de organisme, unele nici măcar direct legate între ele - o consecință a multor evenimente endosimbiotice secundare și chiar terțiare .

Rodopsina microbiană

(2) își schimbă configurația, astfel încât un proton este expulzat din celulă
(3) potențialul chimic face ca protonul să curgă înapoi în celulă
(4) generând astfel energie
(5) în forma de adenozin trifosfat .
Metabolismul fototrofic se bazează pe unul dintre cei trei pigmenți care transformă energia: clorofila , bacterioclorofila și retina . Retina este cromoforul care se găsește în rodopsine . Semnificația clorofilei în conversia energiei luminoase a fost scrisă de zeci de ani, dar fototrofia bazată pe pigmenți retinieni abia începe să fie studiată.
.jpg)
| Videoclip extern | |
|---|---|
|
|
În 2000, o echipă de microbiologi condusă de Edward DeLong a făcut o descoperire crucială în înțelegerea ciclurilor marine de carbon și energie. Au descoperit o genă în mai multe specii de bacterii responsabile de producerea proteinei rodopsină , care până acum nu se mai auzea în bacterii. Aceste proteine găsite în membranele celulare sunt capabile să convertească energia luminii în energie biochimică datorită unei modificări a configurației moleculei de rodopsină pe măsură ce lumina solară o lovește, provocând pomparea unui proton din interior spre exterior și un flux ulterior care generează energia. Rodopsinele de tip arhaeal au fost găsite ulterior printre diferiți taxoni, protiști, precum și în bacterii și arhee, deși sunt rare în organismele multicelulare complexe .
Cercetările din 2019 arată că aceste „bacterii care răpesc soarele” sunt mai răspândite decât se credea anterior și ar putea schimba modul în care oceanele sunt afectate de încălzirea globală. „Descoperirile se desprind de interpretarea tradițională a ecologiei marine găsită în manuale, care afirmă că aproape toată lumina soarelui din ocean este capturată de clorofila din alge. În schimb, bacteriile echipate cu rodopsină funcționează ca mașinile hibride, alimentate cu materie organică atunci când sunt disponibile - ca majoritatea bacteriilor sunt - și de lumina soarelui, atunci când substanțele nutritive sunt rare. "
Există o conjectură astrobiologică numită ipoteza Purple Earth care presupune că formele de viață originale de pe Pământ au fost mai degrabă bazate pe retină decât pe clorofilă, ceea ce ar fi făcut Pământul să pară purpuriu în loc de verde.
Alge marine

| Parte dintr-o serie pe |
| Plancton |
|---|
 |
Algele sunt un termen informal pentru o colecție larg răspândită și diversă de organisme eucariote fotosintetice care nu sunt neapărat strâns legate și sunt astfel polifiletice . Spre deosebire de plantele superioare, algele nu au rădăcini, tulpini sau frunze.
Grupuri de alge
Algele marine au fost plasate în mod tradițional în grupuri precum: alge verzi , alge roșii , alge brune , diatomee , coccolitofori și dinoflagelați .
Alge verzi
Algele verzi trăiesc cea mai mare parte a vieții lor ca celule unice sau sunt filamentoase, în timp ce altele formează colonii formate din lanțuri lungi de celule sau sunt alge marine macroscopice foarte diferențiate. Ei formează un grup informal care conține aproximativ 8.000 de specii recunoscute.
Alge roșii
Algele roșii moderne sunt în mare parte multicelulare cu celule diferențiate și includ multe alge marine notabile . Ca alge coraline , ele joacă un rol important în ecologia recifelor de corali. Ele formează un filum (în litigiu) care conține aproximativ 7.000 de specii recunoscute.

Colonia Cyanidiophyceae , o clasă de alge roșii unicelulare

Algele Porphyra umbilicalis


Alge brune
Algele brune sunt în majoritate multicelulare și includ multe alge marine, inclusiv alge . Ele formează o clasă care conține aproximativ 2.000 de specii recunoscute.
Diatomee

În total, aproximativ 45% din producția primară din oceane este contribuită de diatomee .

Diatomeele sunt unul dintre cele mai frecvente tipuri de fitoplancton

Sunt un grup major de alge care generează aproximativ 20% din producția mondială de oxigen.

Diatomee legate într-un lanț colonial
_Various_diatoms.jpg)



Coccolithophores

Omniprezenta Emiliania huxleyi

Emiliania huxleyi înflorește în sudul Angliei


Coccolithophores sunt aproape exclusiv marine și se găsesc în număr mare în toată zona de lumina soarelui a oceanului . Au plăci de carbonat de calciu (sau solzi) cu funcții incerte numite coccolite , care sunt microfosile importante . Coccolithophores sunt de interes pentru cei care studiază schimbările climatice globale , deoarece odată cu creșterea acidității oceanelor , coccoliturile lor pot deveni și mai importante ca o scufundare de carbon . Cea mai abundentă specie de coccolithophore, Emiliania huxleyi este o componentă omniprezentă a bazei de plancton din rețelele alimentare marine . Sunt folosite strategii de management pentru a preveni inflorirea coccolitoforului legată de eutrofizare, deoarece aceste înfloriri conduc la o scădere a fluxului de nutrienți la niveluri mai scăzute ale oceanului.
Dinoflagelat

Dinoflagelate

Karenia brevis produce maree roșii extrem de toxice pentru oameni


_by_Noctiluca_in_Nagasaki.jpg)
Alge mixotrofe
Alte grupuri

Diplonemidele pot fi abundente în oceanele lumii

În mod tradițional, filogenia microorganismelor, cum ar fi grupurile de alge discutate mai sus, a fost dedusă și taxonomia lor stabilită pe baza studiilor morfologice . Cu toate acestea, evoluțiile filogeneticii moleculare au permis stabilirea relației evolutive a speciilor prin analiza ADN - ului și a secvențelor lor de proteine . Mulți taxoni, inclusiv grupurile de alge discutate mai sus, sunt în proces de reclasificare sau redefinire utilizând filogenetica moleculară. Dezvoltările recente în secvențierea moleculară au permis recuperarea genomilor direct din probele de mediu și evitarea necesității cultivării. Acest lucru a dus, de exemplu, la o extindere rapidă a cunoașterii abundenței și diversității microorganismelor marine . Tehnicile moleculare, cum ar fi metagenomica rezolvată de genom și genomica monocelulară, sunt utilizate în combinație cu tehnici cu randament ridicat .
Între 2009 și 2013, expediția Tara Oceans a traversat oceanele lumii colectând plancton și analizându-le cu tehnici moleculare contemporane. Au găsit o gamă imensă de alge fotosintetice și mixotrofe necunoscute anterior. Printre descoperirile lor s-au numărat diplonemidele . Aceste organisme sunt în general incolore și de formă alungită, de obicei aproximativ 20 µm lungime și cu doi flageli. Dovezile din codul de bare ADN sugerează că diplonemidele pot fi printre cele mai abundente și mai bogate în specii din toate grupurile de eucariote marine.
După mărime
Algele pot fi clasificate după mărime ca microalgă sau macroalgă .
Microalge
Microalgele sunt tipurile microscopice de alge, care nu sunt vizibile cu ochiul liber. Sunt în mare parte specii unicelulare care există ca indivizi sau în lanțuri sau grupuri, deși unele sunt multicelulare . Microalgele sunt componente importante ale protiștilor marini , precum și fitoplanctonul marin . Sunt foarte diverse . S-a estimat că există 200.000-800.000 de specii din care au fost descrise aproximativ 50.000 de specii. În funcție de specie, dimensiunile lor variază de la câțiva micrometri (µm) la câteva sute de micrometri. Sunt special adaptate la un mediu dominat de forțe vâscoase.
- Microalge

Zooxanthellae este o algă fotosintetică care trăiește în interiorul gazdelor precum coralul

Un ciliat unicelular cu zooclorele verzi care trăiesc în interior endosimbiotic

Euglena mutabilis , un flagelat fotosintetic


_(cropped).jpg)
Macroalge

Macroalgele sunt tipurile mai mari, multicelulare și mai vizibile de alge, denumite în mod obișnuit alge marine . Algele marine cresc, de obicei, în apele de coastă puțin adânci, unde sunt ancorate de fundul mării printr-o fermă . Algele marine care devin în derivă se pot spăla pe plaje. Kelp este o alge marine mare maro care formeaza mari subacvatice păduri care acoperă aproximativ 25% din coastele lumii. Acestea se numără printre cele mai productive și dinamice ecosisteme de pe Pământ. Unele alge marine Sargassum sunt planctonice (plutitoare libere) și formează drifturi plutitoare. La fel ca microalga, macroalga (algele marine) sunt din punct de vedere tehnic protiste marine, deoarece nu sunt plante adevărate.


- Macroalge

Alge uriaș este tehnic un protist, deoarece nu este o plantă adevărată, totuși este multicelulară și poate crește până la 50 m

Peștii Sargassum sunt camuflați pentru a trăi printre algele Sargassum în derivă

Această algă cu bule unicelulare trăiește în zone de maree . Poate avea un diametru de 4 cm.




Evoluția plantelor terestre

Un scenariu evolutiv pentru cucerirea terenului de către streptofite
Întâlnirile se bazează aproximativ pe Morris și colab. 2018.
Diagrama din dreapta arată un scenariu evolutiv pentru cucerirea pământului de către streptofite. Algele streptofite includ toate algele verzi și sunt singurele eucariote fotosintetice din care a evoluat flora macroscopică a terenului (linii roșii) . Acestea fiind spuse, de-a lungul evoluției, algele din diferite alte descendențe au colonizat pământul (linii galbene) - dar și algele streptofite au făcut continuu și independent tranziția umedă spre cea uscată (convergența roșu și galben). De-a lungul istoriei, numeroase descendențe au dispărut (etichete X) . Algele terestre de diferite afilieri taxonomice stau pe suprafețele rocilor și formează cruste biologice ale solului . Totuși, din diversitatea algelor stafilofite parafiletice a apărut un organism ai cărui descendenți au cucerit în cele din urmă pământul la scară globală: un organism filamentos - sau chiar parenchimatos - ramificat care a format structuri rizoidale și a experimentat desicarea din când în când. Din această „algă hidro-terestră ipotetică” au apărut descendențele Zygnematophyceae și embriofitelor (plantele terestre). La începuturile sale, traiectoria care ducea la embrioni era reprezentată de cele mai vechi plante terestre - acum dispărute.
Cele mai vechi plante terestre au interacționat probabil cu microbiota substratului benefic , care le-a ajutat la obținerea substanțelor nutritive din substratul lor. Mai mult, primele plante terestre au trebuit să depășească cu succes un baraj de factori de stres terestru (inclusiv lumină ultravioletă și iradiere fotosintetic activă , secetă, schimbări drastice de temperatură etc.). Au reușit deoarece au avut setul potrivit de trăsături - un amestec de adaptări care au fost selectate în strămoșii lor algali hidro-terestri, exaptări și potențialul de co-opțiune a unui set fortuit de gene și căi. Pe parcursul evoluției, unii membri ai populațiilor celor mai vechi plante terestre au dobândit trăsături care sunt adaptive în mediile terestre (cum ar fi o formă de conductanță a apei, structuri asemănătoare stomatelor , embrioni etc.); în cele din urmă, a apărut „ipoteticul ultim strămoș comun al plantelor terestre”. Din acest strămoș, au evoluat briofitele și traheofitele existente . În timp ce repertoriul exact al trăsăturilor ipoteticului ultim strămoș comun al plantelor terestre este incert, acesta va presupune cu siguranță proprietăți ale plantelor vasculare și non-vasculare . Ceea ce este, de asemenea, sigur este că ultimul strămoș comun al plantelor terestre a avut trăsături de ascendență algală.
Plantele marine

Înapoi în Silurian , unele fitoplancton au evoluat în alge roșii , maronii și verzi . Alge verzi au invadat apoi pământul și au început să evolueze în plantele terestre pe care le cunoaștem astăzi. Mai târziu, în Cretacic , unele dintre aceste plante terestre s-au întors la mare ca mangrove și ierburi marine .
Viața plantelor poate înflori în apele salmastre ale estuarelor , unde ar putea crește mangrove sau iarbă de cordon sau iarbă de plajă . Plantele cu flori cresc în adâncimi nisipoase sub formă de pajiști de iarbă de mare , mangrovele se întind pe coastă în regiunile tropicale și subtropicale, iar plantele tolerante la sare prosperă în mlaștinile sărate inundate în mod regulat . Toate aceste habitate sunt capabile să sechestreze cantități mari de carbon și să susțină o gamă biodiversă de animale mai mari și mai mici. Plantele marine pot fi găsite în zonele intertidale și în apele puțin adânci, cum ar fi iarbă de mare , cum ar fi anghila și iarba țestoasă , Thalassia . Aceste plante s-au adaptat la salinitatea ridicată a mediului oceanic.
Lumina este capabilă să pătrundă în primii 200 de metri (660 ft), deci aceasta este singura parte a mării în care plantele pot crește. Straturile de suprafață sunt adesea deficitare în compuși de azot biologic activi. Ciclul azotului marin constă din transformări microbiene complexe care includ fixarea azotului , asimilarea acestuia, nitrificarea , anamox și denitrificarea. Unele dintre aceste procese au loc în apele adânci, astfel încât acolo unde există o revărsare a apelor reci și, de asemenea, în apropierea estuarelor în care sunt prezenți nutrienți de la sol, creșterea plantelor este mai mare. Aceasta înseamnă că zonele cele mai productive, bogate în plancton și, prin urmare, și în pești, sunt în principal de coastă.
Mangrove
Mangrovele oferă habitate importante pentru pepiniere pentru viața marină, acționând ca locuri de ascundere și hrănire pentru forme larvare și juvenile de pești și nevertebrate mai mari. Pe baza datelor prin satelit, suprafața mondială totală a pădurilor de mangrove a fost estimată în 2010 la 134.257 kilometri pătrați (51.837 mile pătrate).



- Spalding, M. (2010) Atlasul mondial de mangrove , Routledge. ISBN 9781849776608 . doi : 10.4324 / 9781849776608 .
Ierburi marine
La fel ca mangrovele, iarbele marine oferă habitate importante de pepinieră pentru formele larvare și juvenile de pești și nevertebrate mai mari. Suprafața mondială totală a pajiștilor cu iarbă de mare este mai dificil de determinat decât pădurile de mangrove, dar a fost estimată în mod conservator în 2003 la 177.000 de kilometri pătrați (68.000 de mile pătrate).

Dragonii de mare camuflați pentru a arăta ca alge marine plutitoare trăiesc în păduri de alge și pajiști de iarbă de mare


| Videoclip extern | |
|---|---|
|
|
Stoichiometrie
Stoichiometria (Măsurarea chimice reactanți și produse ) de producție primară în ocean de suprafață joacă un rol crucial în ciclism de elemente în oceanul global. Raportul dintre elementele de carbon (C), azot (N) și fosfor (P) în materia organică exportată exprimat în termeni de C: N: raportul P ajută la determinarea cât de mult carbon atmosferic este sechestrat în ocean adânc în ceea ce privește disponibilitatea nutrienților limitativi . La scară de timp geologică, raportul N: P reflectă disponibilitatea relativă a nitraților față de fosfat , ambii fiind furnizați extern din atmosferă prin fixarea azotului și / sau continente prin aprovizionarea cu râuri și pierduți prin denitrificare și înmormântare. Pe perioade mai scurte de timp, stoichiometria medie a materiei organice în particule în vrac exportată reflectă stoichiometria elementară a fitoplanctonului , cu influențe suplimentare din diversitatea biologică și procesarea secundară a materiei organice de către zooplancton și bacterii heterotrofe . În fața schimbărilor globale, înțelegerea și cuantificarea mecanismelor care conduc la variabilitatea raporturilor C: N: P sunt cruciale pentru a avea o proiecție exactă a schimbărilor climatice viitoare.
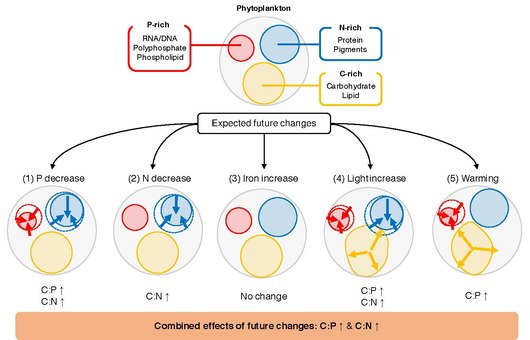
la factorii principali de mediu
O întrebare cheie nerezolvată este ce determină C: N: P de fitoplancton individual. Fitoplanctonul crește în stratul superior luminat de lumină al oceanului , unde cantitatea de nutrienți anorganici, lumina și temperatura variază spațial și temporal. Studiile de laborator arată că aceste fluctuații declanșează răspunsuri la nivel celular, prin care celulele modifică alocarea resurselor pentru a se adapta optim mediului lor ambiental. De exemplu, fitoplanctonul poate modifica alocarea resurselor între aparatul biosintetic bogat în P, aparatul de recoltare a luminii bogat în N și rezervele de stocare a energiei bogate în C. În cadrul unui scenariu tipic de încălzire viitoare, oceanul global este de așteptat să sufere modificări ale disponibilității nutrienților, temperaturii și iradianței . Este posibil ca aceste modificări să aibă efecte profunde asupra fiziologiei fitoplanctonului, iar observațiile arată că speciile de fitoplancton competitive se pot climatiza și se pot adapta la schimbările de temperatură, iradiere și substanțe nutritive pe perioade decenale. Au fost efectuate numeroase experimente de laborator și de teren care studiază relația dintre raportul C: N: P al fitoplanctonului și al factorilor de mediu. Cu toate acestea, este dificil să sintetizăm aceste studii și să generalizăm răspunsul fitoplanctonului C: N: P la modificările factorilor de mediu. Studiile individuale utilizează diferite seturi de analize statistice pentru a caracteriza efectele factorilor de control asupra mediului asupra raporturilor elementare, variind de la un test t simplu la modele mixte mai complexe, ceea ce face ca comparațiile dintre studii să fie dificile. În plus, deoarece modificările caracteristicilor induse de mediu sunt determinate de o combinație de plasticitate ( aclimatizare ), adaptare și istorie de viață, răspunsurile stoechiometrice ale fitoplanctonului pot fi variabile chiar și între speciile strâns legate.
Metaanaliza / revizuirea sistematică este un cadru statistic puternic pentru sintetizarea și integrarea rezultatelor cercetării obținute din studii independente și pentru descoperirea tendințelor generale. Sinteza seminală de Geider și La Roche în 2002, precum și lucrarea mai recentă a lui Persson și colab. în 2010, a arătat că C: P și N: P pot varia cu până la un factor de 20 între celulele pline de nutrienți și celulele limitate de nutrienți. Aceste studii au arătat, de asemenea, că raportul C: N poate fi modest plastic din cauza limitării nutrienților. Un studiu de meta-analiză realizat de Hillebrand și colab. în 2013 a evidențiat importanța ratei de creștere în determinarea stoechiometriei elementare și a arătat că atât raporturile C: P, cât și N: P scad odată cu creșterea ratei de creștere. În 2015, Yvon-Durocher și colab. a investigat rolul temperaturii în modularea C: N: P. Deși setul lor de date a fost limitat la studii efectuate înainte de 1996, au arătat o relație semnificativă statistic între C: P și creșterea temperaturii. MacIntyre și colab. (2002) și Thrane și colab. (2016) au arătat că iradianța joacă un rol important în controlul raporturilor C: N și N: P celulare optime. Cel mai recent, Moreno și Martiny (2018) au furnizat un rezumat cuprinzător al modului în care condițiile de mediu reglează stoichiometria celulară dintr-o perspectivă fiziologică.
Stoichiometria elementară a fitoplanctonului marin joacă un rol critic în ciclurile biogeochimice globale prin impactul său asupra ciclului nutrienților, producției secundare și exportului de carbon. Deși au fost efectuate experimente de laborator ample de-a lungul anilor pentru a evalua influența diferiților factori de mediu asupra compoziției elementare a fitoplanctonului, lipsește încă o evaluare cantitativă cuprinzătoare a proceselor. Aici, răspunsurile raporturilor P: C și N: C ale fitoplanctonului marin au fost sintetizate la cinci factori principali (fosfor anorganic, azot anorganic, fier anorganic, iradiere și temperatură) printr-o meta-analiză a datelor experimentale din 366 de experimente din 104 articole de revistă. Aceste rezultate arată că răspunsul acestor rapoarte la modificările macronutrienților este consecvent în toate studiile, unde creșterea disponibilității nutrienților este legată pozitiv de modificările raporturilor P: C și N: C. Rezultatele arată că fitoplanctonul eucariot este mai sensibil la modificările macronutrienților în comparație cu procariotele, posibil datorită dimensiunii celulare mai mari și abilităților lor de a-și regla rapid modelele de expresie genică. Efectul iradianței a fost semnificativ și constant în toate studiile, unde o creștere a iradianței a scăzut atât P: C, cât și N: C. Raportul P: C a scăzut semnificativ odată cu încălzirea, dar răspunsul la schimbările de temperatură a fost mixt în funcție de modul de creștere a culturii și de faza de creștere în momentul recoltării. Împreună cu alte condiții oceanografice ale girurilor subtropicale (de exemplu, disponibilitate scăzută a macronutrienților), temperatura ridicată poate explica de ce P: C este constant scăzut în oceanele subtropicale. Adăugarea de fier nu a schimbat în mod sistematic nici P: C, nici N: C.
Cronologie evolutivă
.png)

Vezi si
- Alge
- Plante acvatice
- Istoria evolutivă a plantelor
- Evoluția plantelor
- Cronologia evoluției plantelor
- Evoluția fotosintezei
Referințe
Lecturi suplimentare
- Falkowski, Paul (Ed.) (2013) Productivitatea primară în Sea Springer. ISBN 9781468438901 .
- Falkowski, Paul și Raven, John A. (2013) Photosynthesis Aquatic Ediția a doua revizuită, Princeton University Press. ISBN 9781400849727 .
- Falkowski P și Knoll AH (2011) Evoluția producătorilor primari în Sea Academic Press. ISBN 9780080550510 .
- Kirk, John TO (2010) Light and Photosynthesis in Aquatic Ecosystems Ediția a treia revizuită, Cambridge University Press. ISBN 9781139493918 .